Farzana Y Zaman
Fifth Year Medicine/BMedSci (Undergraduate)
Monash University
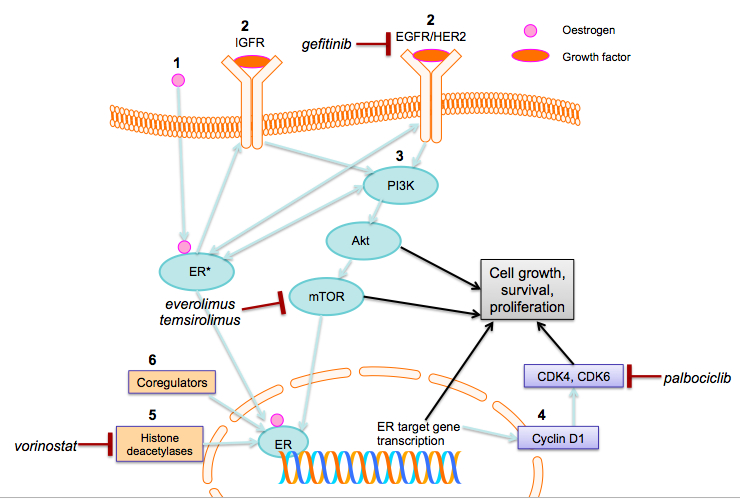
Farzana is currently doing a BMedSci in the area of breast cancer research. She hopes to one day be an oncologist and clinician-scientist, and is looking forward to her final year of medical school in 2016. She plays violin in the Monash Medical Orchestra and enjoys devouring novels, travelling and spending time with her cat.