Dr. Jazlyn Read
BPhty, MBBS
Intern, Royal Brisbane and Women’s Hospital
Jazlyn is an intern at the Royal Brisbane and Women’s Hospital, and wrote this article while studying final-year medicine at Griffith University. In her spare time she enjoys travelling and languages.
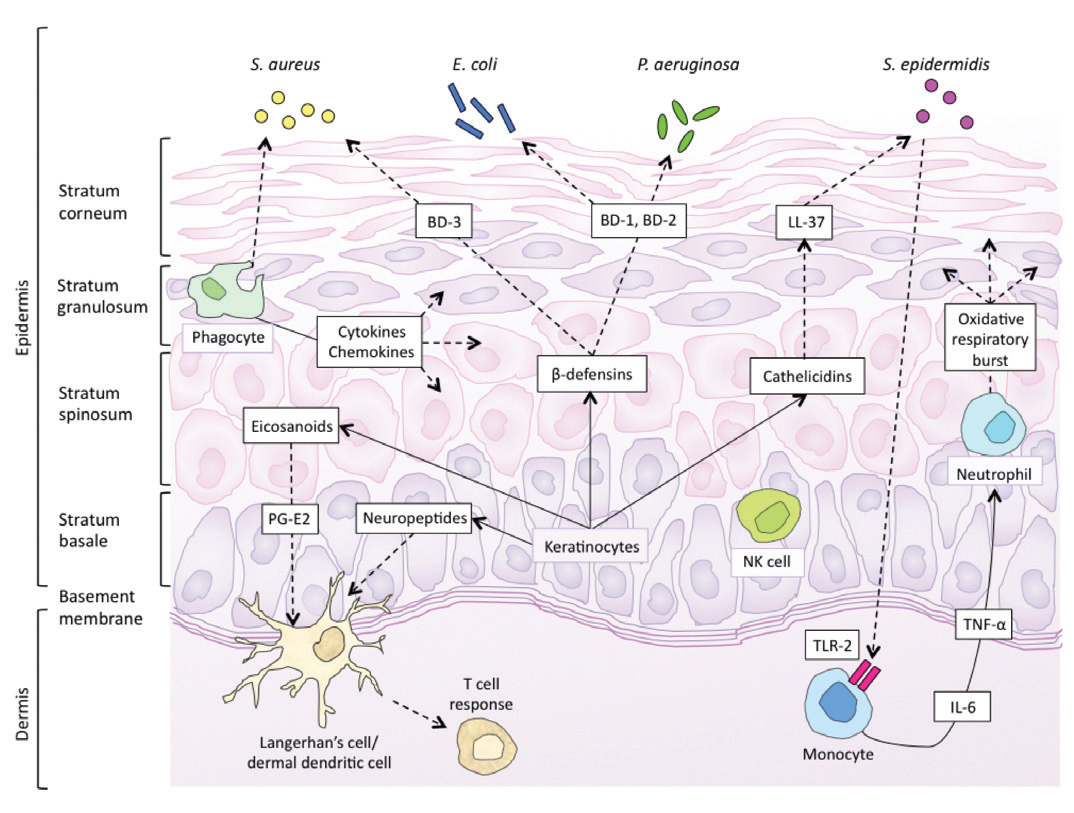
 Skin forms a dynamic interface with the external environment and is a complex organisation of cell types and associated structures that performs many essential functions. Although the stratum corneum of full-term neonates is analogous to that of adult skin, structural and compositional differences of the skin renders the newborn more susceptible to bacterial colonisation. Particularly for the preterm neonate, impaired cutaneous barrier function and an immature immune system reduce the capacity to defend against bacterial pathogens. The majority of cutaneous bacterial infections are localised to the skin and are easily treated, however, systemic bacterial infection and disseminated disease in the neonatal period may be life-threatening.
Skin forms a dynamic interface with the external environment and is a complex organisation of cell types and associated structures that performs many essential functions. Although the stratum corneum of full-term neonates is analogous to that of adult skin, structural and compositional differences of the skin renders the newborn more susceptible to bacterial colonisation. Particularly for the preterm neonate, impaired cutaneous barrier function and an immature immune system reduce the capacity to defend against bacterial pathogens. The majority of cutaneous bacterial infections are localised to the skin and are easily treated, however, systemic bacterial infection and disseminated disease in the neonatal period may be life-threatening.![Table 1. Differences between newborn and adult skin. [1,2]](http://www.amsj.org/wp-content/uploads/2015/04/v5_i1_a22_1.jpg)