Justin Smith
4th Year Undergraduate MBBS Student, James Cook University
4th Year Undergraduate MBBS Student, James Cook University
Justin Smith is a fourth year medical student at James Cook University.
Thursday, August 31st, 2017
Aim: To ascertain the function of miRNA-34a in hepatocellular carcinoma (HCC) and to assess its use as a therapeutic agent through the analysis of pre-clinical and clinical trials.
Discussion: Multiple studies found that miRNA-34a was down-regulated in the majority of human HCC samples and subsequently had a tumour suppressor role via the inhibition of a number of target genes essential for carcinogenesis. MRX34, a miRNA-34a mimic, is currently in an ongoing phase I clinical trial. Interim data has indicated that this therapy has a manageable safety profile, with a partial response observed in one patient. The combination of miRNA-34a with other agents has also proven to exert enhanced anti-tumour effects. Conversely, many studies have reported that miRNA-34a was up-regulated in HCC samples, particularly in those with activation of the beta-catenin pathway.
Conclusion: Pre-clinical studies have shown promising results in the use of a miRNA-34a mimic in HCC as a single agent or as a combination therapy, however, the results from the phase I trial are yet to be fully established. The mechanisms of miRNA-34a in HCC remain to be elucidated, with further research required into its proposed oncogenic role, especially relating to the clinical implications of this interaction.
Hepatocellular carcinoma (HCC) is the second most common cause of cancer-related deaths worldwide and is the most common form of liver cancer, accounting for between 85% and 90% of primary liver cancers [1]. The major risk factors for hepatocellular carcinoma include hepatitis B (HBV) or C virus (HCV) infection, smoking, alcohol, and aflatoxin [2]. Currently the prognosis for HCC is poor with Australian statistics indicating that the five year relative survival rate for primary liver cancers is only 16% [3]. The non-specific tyrosine kinase inhibitor sorafenib currently represents the only effective treatment against HCC [4]. This poor prognostic outlook and the limited availability of targeted molecular agents for HCC has led to the development of new therapies such as microRNAs (miRNAs).
miRNAs are short (19-24 nucleotides), non-coding RNA molecules that are post-transcriptional regulators of gene expression. Initially, miRNA is transcribed as primary miRNA (pri-miRNA), which is processed into precursor miRNA (pre-miRNA). This is then transported from the nucleus into the cytoplasm where it is processed into its mature form by the enzyme Dicer. The mature miRNA forms part of the RNA-induced silencing complex (RISC), which is responsible for regulating the output of protein-coding genes. These miRNAs interact with the 3’ untranslated regions (UTRs) of the protein-coding genes to result in a decrease in protein output via mRNA degradation or translational repression [5]. Alternatively, miRNAs can result in post-transcriptional stimulation of gene expression via a multitude of direct and indirect mechanisms [6]. It is estimated that a single miRNA can target hundreds to over one thousand different mRNAs, ultimately resulting in miRNAs being responsible for the regulation of around 20-30% of all protein-encoding genes [7]. These miRNAs have been reported to have key roles in cancer initiation, progression, and metastasis [8]. Oncogenic miRNAs are miRNAs that are up-regulated in cancer cells and promote carcinogenesis via the inhibition of tumour suppressor genes. Conversely, the miRNAs that are decreased in cancer cells are known as tumour suppressor miRNAs, as they normally inhibit proto-oncogenes to prevent cancer from developing [9]. The mammalian miRNA-34 family consists of miRNA-34a, which is encoded via its own individual transcript, and miRNA-34b and miRNA-34c, which possess a common primary transcript [10]. Due to the promising and extensive research conducted on miRNA-34a, this review article focused specifically on this particular isoform. Dysregulation of miRNA-34a has been implicated in a wide variety of cancers, including prostate, colon [11], and HCC [12]. The purpose of this review is to analyse the specific role of miRNA-34a in HCC, including addressing contradictory findings and investigating the recent clinical trials.
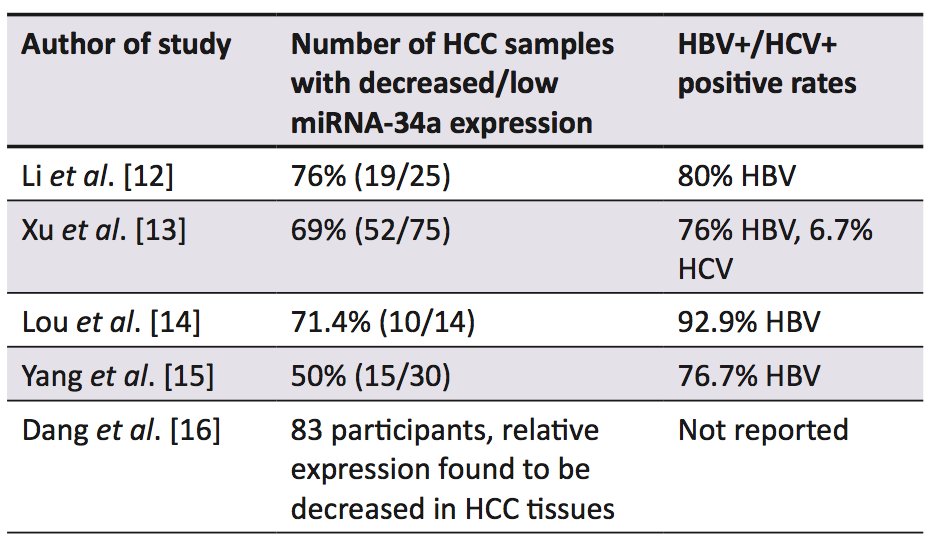
A range of studies established that in the majority of human HCC samples, miRNA-34a expression was decreased in comparison to the surrounding non-cancerous liver tissue (Table 1) [12-16]. A murine model of hepatocarcinogenesis induced by a methyl-deficient diet also resulted in the down-regulation of miRNA-34a [17]. Low expression of miRNA-34a in HCC samples has been correlated with a shorter overall [13-15] and disease-free survival [14], as well as higher recurrence rates [13] when compared with samples that displayed up-regulation of miRNA-34a. The decreased expression of miRNA-34a is thought to be caused by genetic alterations such as deletions, point mutations, or chromosomal translocations of its genomic region 1p36 [18], which is common in HCC [19, 20]. Alternatively, this decreased expression has been linked to inactivating mutations of the p53 gene [18], as the induction of miRNA-34a is correlated with p53 status [21]. Furthermore, epigenetic silencing of miRNA-34a has been implicated with these decreased expression levels in multiple forms of cancer, via abnormal CpG methylation in its promoter region [22].
The administration of a miRNA-34a mimic (MRX34) has been shown to cause inhibition of a number of genes within multiple oncogenic pathways such as Wnt/ beta-catenin, c-MET, VEGF, hedgehog, and MAPK (all of which have been implicated in hepatocarcinogenesis), as well as stimulating multiple genes of the p53 pathway [23]. Daige et al. explored a diverse range of HCC related pathways, demonstrating how miRNA-34a exerts its anti-cancer effects by modulation of a number of genes responsible for processes such as metastasis, cellular proliferation, cell cycle regulation, apoptosis, and cellular senescence [23].
A number of cell culture studies investigated the effects of ectopic expression of miRNA-34a in the HepG2 cell line, with contradicting results [12,16,18]. Ectopic expression of miRNA-34a caused significant inhibition of cellular proliferation at 72 hours [18] and 96 hours [16] post- transfection. In addition, miRNA-34a was demonstrated to regulate the cell cycle via inducing G1 arrest [18]. Furthermore, it has been found that miRNA-34a can induce apoptosis, as determined by increased caspase 3/7 activity [16]. In contrast, other reports claimed that there was no effect on cellular proliferation, G1 arrest, or apoptosis, [12,18], highlighting the conflicting information within the current literature. The discrepancies between these three studies could partially be explained by the varying methods used to express miRNA-34a, and the different measurement times post-transfection (48, 72 or 96 hours) [16]. Additional research has found that miRNA-34a induces apoptosis in HCC cells via binding to the 3’ UTR of the Bcl-2 mRNA, causing inhibition of its translation [15]. Over-expression of miRNA-34a has also been correlated with a decreased expression of Bcl-2 in a number of other HCC studies [14,24].
Down-regulation of miRNA-34a expression has been associated with metastatic HCC [12,16,25]. Multiple studies have established that the ectopic[1] expression of miRNA-34a in the HepG2 cell line inhibits tumour cell migration and invasion via silencing of the c-Met gene, which subsequently decreases the c-Met-induced phosphorylation of extracellular signal-regulated kinases 1 and 2 (ERK1/2) [12,16]. Cheng et al. also proposed that miRNA-34a prevents tumour migration, invasion, and metastasis by decreasing cathepsin D [18], a protease that contributes to the degradation of the basement membrane [26]. Furthermore in the HCC cell line Hca-F (high lymphatic metastatic potential) it was determined through in vitro and in vivo studies in mice that the ectopic expression of miRNA-34a caused a reduction in the metastatic potential [27].
Earlier studies showed that miRNA-34a induced cellular senescence via cell cycle arrest in pathways that were telomere-independent [13, 28]. Recently, miRNA-34a over-expression has been shown to induce senescence in HCC cells in a telomere-dependent manner, regulated by p53. This cellular senescence occurs by the inhibition of FoxM1 and c-Myc, which causes the inactivation of telomerase activity, resulting in telomere shortening (Figure 1) [13].

There is a vast array of delivery systems, both viral and non-viral, that are used to increase miRNA expression. Both mechanisms are associated with advantages and disadvantages [29]. Rubone and MRX34 are two prominent non-viral delivery methods that have been investigated in HCC studies [23,24]. Rubone is a small molecular modulator of miRNA-34a that was shown to induce miRNA-34a expression selectively in HCC cells (although only those with wild-type or mutant p53, not with p53 deletion), causing inhibition of tumour growth both in vitro and in vivo in the HepG2 xenograft mouse model. Xiao et al. also found that this miRNA-34a modulator displayed similar or even greater anti-HCC activity than sorafenib, the current treatment for advanced HCC [24]. MRX34 is a double-stranded miRNA-34a mimic that is delivered by liposomes [30]. The systemic delivery of this molecule resulted in tumour regression during in vivo studies in two different xenograft mouse models (Hep3B and HuH7) of liver cancer [23]. An oncolytic adenoviral vector that co-expressed miRNA-34a and IL-24 has also been studied in a HCC model. This was found to cause increased anti-tumour activity both in vitro and in vivo, predominantly via the downregulation of SIRT1 and Bcl-2 [14].
Currently there is an ongoing phase I trial of MRX34 [31], which commenced in April 2013 and was originally indicated for patients with primary liver cancer or cancers with metastasis to the liver [32]. It was then gradually expanded to include patients with other advanced solid tumours (with or without liver metastasis) and haematological malignancies (lymphoma and multiple myeloma) [31]. There were 75 patients with advanced solid tumours enrolled in this study and 30 of these had HCC [33]. This trial’s data revealed that partial responses to the treatment, as per the RECIST guidelines (Table 2) [34], were observed in one patient with HCC, one with melanoma, [33] and one with advanced renal cell carcinoma [35]. Furthermore a number of patients in this trial were found to have attained a stable disease state [35]. This interim phase I data also determined that MRX34 has a manageable safety profile [33]. A recent press release from MIRNA therapeutics has indicated that phase II clinical trials will commence by the end of 2016 and will consist of two studies, one on renal cell carcinoma and the other on melanoma [35]. In terms of HCC-specific trials with MRX34, the future direction is currently unclear and with limited data available at the present time it is difficult to draw any definitive conclusions.

Yang et al. demonstrated that ectopic expression of miRNA-34a resulted in the sensitisation of HCC cells to sorafenib-induced apoptosis and toxicity via inhibiting expression of Bcl-2 [15]. Additionally, the administration of miRNA-34a was found to sensitise HCC cells to chemotherapy (cisplatin) in vitro through the AXL pathway [36]. The combination of a miRNA-34a mimic and C-met inhibitor also resulted in a greater inhibition of cell growth and induction of apoptosis in vitro than either of these two therapies alone [16]. However, to establish more definitive results, further research is required in this field, particularly in regards to clinical trials.
Conversely, a number of other studies have shown increased expression of miRNA-34a in both murine and human HCC tissues [37-41], suggesting it may have an oncogenic role in addition to its tumour suppressor role. A recent article has investigated these claims and found that miRNA-34a displays oncogenic properties in liver tumours with beta-catenin activation [40]. Increased beta-catenin activation is most commonly caused by mutations in the CTNNB1 gene (the gene encoding beta-catenin), and this is estimated to occur in 20-40% of HCCs [42]. Gougelet and colleagues demonstrated using ApcKO mice (ApcKO causes activation of the Wnt/beta-catenin pathway [43]) that administration of a miRNA-34a inhibitor (LNA-34a) caused increased expression of hepatocyte nuclear factor 4a. This leads to increased apoptosis predominantly via caspase 2 activation and decreased cell proliferation through inhibition of cyclin D1 (Figure 2). This theory was then postulated to complement the data of the studies listed in Table 1, by accounting for those samples that showed up-regulation of miRNA-34a expression. The disparity in results between studies is thought to be due to the varying causes of HCC [40]. For example, the majority of HCC samples from the studies in the Table 1 were HBV+, and this was found to be associated with a lower frequency of CTNNB1 mutations [44]. Conversely HCC resulting from HCV infection has been shown to have a higher rate of CTNNB1 mutations [45]. However, these findings contradict the theory that miRNA-34a has a tumour suppressor function, and with relatively limited research on this oncogenic pathway, further investigation is required. Studies investigating the concept of miRNA-34a having a tumour suppressor or oncogenic function depending on the cause of the tumour would also be important, as well as an investigation of the clinical implications of this relationship.

A number of limitations were identified within this review, particularly relating to the conflicting information and the limited availability of clinical trial results. Contradictory information was noted on a number of occasions, especially with the use of a miRNA-34a inhibitor for HCC, making it difficult to evaluate a clear clinical benefit to this potential therapy. The data surrounding the clinical trial was also restricted as the trial is ongoing. Subsequently, all the data had to be sourced from press releases and abstracts from presentations at conferences, which were all funded by MIRNA therapeutics; thus a potential conflict of interest was noted.
Results have indicated that miRNA-34a has a tumour suppressor function in HCC and is responsible for the down-regulation of a number of genes involved in carcinogenesis. There is, however, contradicting information described in studies investigating these parameters, highlighting the complexity of this topic. Rubone and MRX34 are two prominent miRNA-34a delivery systems that were shown to exert anti-tumour activity in pre-clinical models. Additionally, MRX34 has been commenced in a clinical phase I trial that is still currently ongoing, with a partial response already observed in one patient. However, the future of HCC-specific MRX34 trials remains unclear as limited information is currently available. Based on the promising results of miRNA-34a as a combination therapy, this is an area that requires further investigation through clinical trials. Conversely, other sources have found that miRNA-34a plays an oncogenic role in HCC, particularly in those with beta-catenin activation. Subsequently, it was demonstrated that miRNA-34a inhibitors should be used in these instances. Further research is necessary in order to ascertain the clinical implications of using a miRNA-34a mimic or inhibitor depending on the beta-catenin mutation status of the patient.
None declared.